
Козулько Г.А., Козулько Т.Н.
Государственный Национальный парк "Беловежская пуща"
Научные исследования в Беловежской пуще начались около ста лет назад, тем не менее почвенные беспозвоночные, являющиеся одним из наиболее многочисленных и важных компонентов лесного биогеоценоза, остаются наименее изученным зоологическим объектом (Karpinski, Okolow, 1969; Okolow, 1976, 1983; Ковальков, Балюк, Будниченко, 1985). До сих пор неизвестны структурная организация и распределение сообществ беспозвоночных в основных типах леса пущи, недостаточно изучены их состав и плотность. По некоторым группам почвенных животных отсутствуют инвентаризационные списки. В связи с вышеизложенным основная цель настоящей работы - изучить состав, плотность, зоомассу и распределение сообществ почвенных беспозвоночных (мезофауны) в основных типах леса Беловежской пущи.
Почвенную мезофауну изучали в лесах Беловежской пущи в 1988 - 94 гг. на 17 постоянных пробных площадях, заложенных в наиболее распространенных в пуще типах леса (табл. 1). Изучение почвенной мезофауны проводили 4 раза в год (апрель, июнь, август, октябрь). В каждом биотопе за один раз (в течение месяца) брали 16 проб, за полевой сезон - 64 (4 м2). Всего за время исследований взято 1 120 пробы, собрано 24 550 экз. беспозвоночных животных. Среди беспозвоночных выделены 3 группы (дождевые черви, жуки-щелкуны, двукрылые), являющиеся хорошими индикаторами почвенных условий и использованные для диагностики почвенных процессов, которые определяли до вида.
Таблица 1
Лесоводственно-таксационная характеристика пробных площадей
| № | Тип леса | Квар-тал | Состав | Воз-раст, лет | Бо-ни-тет | Пол-нота | Запас древе-сины, м3\га |
Под-ле-сок | Под-рост | Тип почвы | |
| 1 ярус | 2 ярус | ||||||||||
| 1. | С.лиш. | 989 А | 10С | 65 | IY | 0,5 | 90 | мож. | нет | Дерново-слабоподзолистая песчаная подстилаемая песком рыхлым | |
| 2. | С.мш. | 826 А | 10С ед. Б | 60 | I | 0,8 | 230 | нет | нет | Дерново-слабоподзолистая песчаная подстилаемая песком рыхлым | |
| 3. | С.мш. | 826 Г | 10С ед. Б | 10Е ед. Б | 155 | II | 0,7 | 310 | нет | нет | Дерново-слабоподзолистая песчаная подстилаемая песком рыхлым |
| 4. | С.ел.-кис. | 827 А | 9СIЕ ед. Б | 9ЕIГ ед. Д | 185 | I | 0,7 | 410 | нет | ель | Дерново-слабоподзолистая песчаная подстилаемая супесью рыхлой |
| 5. | С.граб. -кис. |
830 Г | 7С3Е ед. Б,Д | 7Г3Е ед. Б,Д | 210 | I | 0,7 | 712 | нет | нет | Дерново-слабоподзолистая песчаная подстилаемая супесью рыхлой |
| 6. | С.чер. | 942 Б | 9СIБ | 70 | I | 0,7 | 230 | круш. | нет | Дерново-среднеподзолистая песчаная подстилаемая песком рыхлым | |
| 7. | С.чер. | 708 Д | 8С2Е ед. Б | 10Е | 155 | I | 0,8 | 626 | нет | ель | Дерново-слабоподзолистая песчаная подстилаемая моренным суглинком |
| 8. | С.долг. | 942 Б | 9СIБ | 70 | III | 0,7 | 220 | круш. | ель | Торфяно-глеевая на оглеенном песке рыхлом | |
| 9. | Е.мш. | 479 Б | 8ЕIСIБ ед. Ос | 8ЕIСIБ ед. Г,Д | 100 | I | 0,7 | 398 | нет | ель | Дерново-слабоподзолистая песчаная подстилаемая песком рыхлым |
| 10. | Е.кис. | 742 А | 8Е2С ед. Д,Ос,Б | 10Е ед. Д,Б,Ос,Г | 110 | I | 0,7 | 522 | нет | ель | Дерново-слабоподзолистая супесчаная подстилаемая песком связным |
| 11. | Е.чер. | 829 Г | 8ЕIСIД | 8Е20С ед. Д | 140 | II | 0,8 | 386 | нет | ель | Дерново-среднеподзолистая песчаная подстилаемая моренным суглинком |
| 12. | Д.кис.(скал.) | 807 А | 10Д ед. Кл. | 10Гед. Е,Кл | 175 | II | 1,0 | 453 | нет | нет | Бурая лесная оподзоленная супесчаная подстилаемая моренным суглинком |
| 13. | Д.кис.(чер.) | 741 А | 8Д2Е ед. Кл,Б | 9Г1Е ед. Кл,Лп | 165 | II | 0,8 | 377 | нет | нет | Бурая лесная псевдоподзолистая супесчаная подстилаемая моренным суглинком |
| 14. | Г.кис. | 863 В | 6Г4Е+Ос ед. Кл,Б,Д | 9Г1Е ед. Кл | 115 | I | 0,8 | 407 | нет | нет | Дерново-слабоподзолистая песчаная подстилаемая моренным суглинком |
| 15. | Я.кис. | 777 В | 4Я1Д4Е1Ол | 9Г1Е | 140 | II | 0,8 | 332 | нет | нет | Дерново-слабоподзолистая супесчаная подстилаемая моренным суглинком |
| 16. | Б.ел.-кис. | 711 Б | 7Б1С1Ос1Лп ед. Е,Д | 9Е1Д ед. Г | 50 | I | 0,8 | 284 | нет | ель | Дерново-слабоподзолистая супесчаная подстилаемая моренным суглинком |
| 17. | Б.граб. -кис. |
908 В | 8Б2Ос ед. Г,Д | 5Г2Е1Кл1Д1Е ед. Ос | 50 | I | 0,9 | 346 | нет | нет | Дерново-слабоподзолистая песчаная подстилаемая моренным суглинком |
Примечание: 1 - Сосняк лишайниковый; 2 - С. мшистый (60 лет); 3 - С. мшистый (155 лет); 4 - С. елово-кисличный; 5 - С. грабово-кисличный; 6 - С. черничный (70 лет); 7 - С. черничный (155 лет); 8 - С. долгомошный; 9 - Ельник мшистый; 10 - Е. кисличный; 11 - Е. черничный; 12 - Дубрава кисличная (дуб скальный); 13 - Д. кисличная (дуб черешчатый); 14 - Грабняк кисличный; 15 - Ясенник кисличный; 16 - Березняк елово-кисличный; 17 - Б. грабово-кисличный
При учетах беспозвоночных животных применяли метод почвенных раскопок с ручной разборкой проб (Гиляров, 1941). Пробы брали площадью 25 см Х 25 см (0,0625 м2) на глубину встречаемости почвенных животных (до 40 см). Разборку подстилки осуществляли несколько усовершенствованной методикой: просеянную мелкую фракцию подстилки собирали в полиэтиленовые мешочки, доставляли их в лабораторию и разбирали под электрической лампой мощностью 200 Вт (это дало возможность наиболее полно учесть мелкие экземпляры пауков, стафилинид, личинок жуков и других мелких беспозвоночных мезофауны).
Статистическая обработка материала проведена по общепринятым формулам и методикам (Зайцев, 1984; Количественные методы ..., 1987). Сравнительный анализ группировок почвенных беспозвоночных проводили по методу Маунтфорда на основе коэффициентов Жаккара [1] и Наумова [2] (Чернов, 1975):
| [1] | Ks = | С | ; |
| —————— | |||
| A + B - C |
| [2] | Kn = | ΣCmin - 100 | ; |
| ————————— | |||
| A + B - ΣCmin |
где С - число видов, общих для двух сравниваемых группировок; А - число видов в первой группировке; В - число видов во второй группировке.
Проверка правильности определения видовой принадлежности личинок щелкунов проведена Л. Пеневым (Болгария), имаго щелкунов - О.Р. Александровичем (БГПУ, Беларусь). Правильность определения личинок двукрылых проверена по эталонным коллекциям, хранящимся в Институте Проблем Экологии и Эволюции (г. Москва). Считаю своим приятным долгом выразить признательность специалистам, оказавшим помощь в определении материала.
В сосняках Беловежской пущи установлена высокая плотность почвенных беспозвоночных. В наиболее сухом типе - сосняке лишайниковом, отмечена наименьшая плотность почвенной мезофауны (243,3 экз/м2) (табл. 2, 3). Наиболее плотно заселены почвы сосняка долгомошного - 585,5 экз/м2. Наименьшая зоомасса беспозвоночных характерна для сосняка лишайникового - 1,9 г/м2 (рис. 1). Максимальная общая зоомасса почвенной мезофауны зарегистрирована в сосняках кисличных - 4,79 г/м2 (рис. 2). В средневозрастных монодоминантных сосняках с упрощенной структурой фитоценоза прослеживается зависимость возрастания общей плотности и зоомассы беспозвоночных с увеличением влажности и богатства почвы. В старовозрастных многоярусных сосняках данные закономерности проявляются сложнее.
Таблица 2
Средняя плотность (экз/м2) почвенных беспозвоночных средневозрастных сосняков
| Группы беспозвоночных |
Сосняк лишайниковый |
Сосняк мшистый |
Сосняк черничный |
Сосняк долгомошный |
| Lumbricidae | 3,0±0,9 | 8,5±2,0 | 10,0±2,1 | 5,8±1,1 |
| Mollusca | – | + | 2,2±0,8 | 11,2±1,9 |
| Oniscoidea | – | – | 1,2±0,6 | 3,5±1,1 |
| Arachnida | 93,8±7,6119,0±8,8 | 152,3±8,6 | 330,5±15,4 | |
| Chilopoda | 2,0±0,8 | 6,8±1,4 | 10,5±1,9 | 11,2±1,8 |
| Diplopoda | – | – | 19,0±4,3 | 92,5±8,2 |
| Insecta | 144,5±13,3 | 149,8±9,8 | 226,8±11,4 | 130,8±7,3 |
| Hemiptera | 6,2±1,5 | 3,0±1,0 | 15,0±2,2 | 4,2±1,4 |
| Dermaptera | – | + | – | + |
| Psocoptera | 10,8±3,2 | + | 1,7±0,7 | – |
| Neuroptera | + | – | 1,8±0,7 | + |
| Rhaphidioptera | 3,0±1,1 | + | + | – |
| Homoptera | + | 2,2±0,9 | 2,0±0,7 | 33,5±4,6 |
| Blattoptera | – | 1,0±0,5 | 3,7±1,8 | + |
| Mecoptera | + | – | – | – |
| Coleoptera | 104,3±9,5 | 124,3±8,1 | 164,8±8,4 | 72,8±4,1 |
| Carabidae | 13,0±2,3 | 7,2±1,5 | 5,2±1,1 | 8,8±1,5 |
| Staphylinidae | 63,5±9,4 | 48,0±5,2 | 104,6±6,3 | 43,5±3,4 |
| Elateridae | 4,8±1,5 | 43,8±4,4 | 38,5±4,1 | 9,2±1,7 |
| Curculionidae | 11,8±2,3 | 22,0±3,0 | 5,5±1,4 | 2,3±0,7 |
| Scarabaeidae | + | + | 1,5±0,6 | – |
| Прочие Coleoptera | 10,7±2,0 | 2,5±0,8 | 9,5±1,7 | 9,0±1,4 |
| Lepidoptera | + | 6,8±1,9 | 7,0±1,5 | 7,2±1,3 |
| Hymenoptera | 1,3±0,6 | 1,2±0,6 | 6,8±1,2 | 3,5±0,8 |
| Diptera | 17,2±2,6 | 9,8±2,0 | 23,8±4,8 | 8,5±1,3 |
| Общая плотность | 243,3±18,2 | 284,3±15,1 | 422,0±16,6 | 585,5±22,2 |
Примечание: здесь и далее + - единично встречаемые особи, – - отсутствие особей
Таблица 3
Средняя плотность (экз/м2) почвенных беспозвоночных старовозрастных сосняков
| Группы беспозвоночных |
Сосняк мшистый |
Сосняк елово-кисличный |
Сосняк грабово-кисличный |
Сосняк черничный |
| Lumbricidae | 3,5±0,9 | 12,2±2,2 | 9,5±1,5 | 7,5±2,0 |
| Mollusca | + | 1,0±0,4 | + | 1,5±0,6 |
| Arachnida | 127,5±14,1 | 77,8±5,8 | 122,0±8,2 | 136,5±11,5 |
| Chilopoda | 3,5±0,9 | 6,4±1,8 | 5,3±1,2 | 11,2±2,4 |
| Diplopoda | – | + | 3,7±1,2 | 2,5±0,9 |
| Insecta | 147,2±8,1 | 193,0±10,1 | 229,2±14,1 | 199,8±10,0 |
| Hemiptera | 6,0±1,6 | 26,6±3,5 | 4,0±1,4 | 3,5±1,2 |
| Dermaptera | 1,0±0,5 | 3,8±2,1 | 4,3±2,0 | 2,0±0,9 |
| Psocoptera | 2,7±1,5 | 1,0±0,5 | 1,5±0,7 | + |
| Neuroptera | – | + | + | – |
| Rhaphidioptera | 3,8±1,2 | – | – | – |
| Homoptera | 1,2±0,8 | 12,0±2,9 | 3,5±1,2 | + |
| Blattoptera | 3,5±1,0 | + | 1,2±0,6 | 2,5±1,1 |
| Mecoptera | – | + | – | – |
| Coleoptera | 109,0±7,5 | 115,4±7,8 | 163,8±12,5 | 161,0±9,9 |
| Carabidae | 5,0±1,3 | 9,8±1,4 | 8,8±1,3 | 9,0±1,9 |
| Staphylinidae | 46,2±5,0 | 69,2±5,3 | 99,2±10,8 | 92,2±8,1 |
| Elateridae | 41,5±4,7 | 17,2±2,3 | 30,0±3,7 | 45,0±4,7 |
| Curculionidae | 7,0±1,3 | 14,2±3,4 | 5,5±1,5 | 9,3±1,7 |
| Scarabaeidae | 2,0±0,9 | 1,6±0,6 | 11,3±2,5 | – |
| Прочие Coleoptera | 7,3±1,6 | 3,4±0,8 | 9,0±1,4 | 5,5±1,1 |
| Lepidoptera | 3,5±1,2 | 3,0±0,8 | 1,3±0,5 | 8,7±1,8 |
| Hymenoptera | 1,7±0,6 | 3,6±0,9 | 3,5±1,2 | 2,0±0,7 |
| Diptera | 14,8±2,0 | 26,4±3,6 | 45,8±4,7 | 18,8±2,8 |
| Общая плотность | 282,0±17,9 | 290,6±14,0 | 370,0±18,1 | 359,0±17,8 |
Примечание: обозначения смотри в таблице 2
 |
| Рис. 1. Зоомасса (г/м2) основных групп почвенных беспозвоночных в cредневозрастных сосняках |
Примечание: 1 - общая; 2 - дождевые черви; 3 - моллюски; 4 - мокрицы; 5 - паукообразные; 6 - губоногие многоножки; 7 - двупарноногие многоножки; 8 - насекомые; 9 - жесткокрылые; 10 - двукрылые;
сокращения типов леса смотри в таблице 1
 |
| Рис. 2. Зоомасса (г/м2) основных групп почвенных беспозвоночных в cтаровозрастных сосняках |
Примечание: обозначения смотри на рисунке 1
В сосняках среди основных групп беспозвоночных массовыми являются насекомые (130,8 - 229,2 экз/м2) и паукообразные (77,8 - 330,5 экз/м2) (табл. 2, 3), что составляет соответственно 22,3 - 66,4% и 26,8 - 56,5% от общей плотности беспозвоночных (рис. 3). В сосняке долгомошном преобладают паукообразные (330,5 экз/м2; 56,5%), а плотность насекомых несколько ниже (130,8 экз/м2; 22,3%). Из насекомых доминируют жесткокрылые (55,6 - 83,0%) и двукрылые (6,5 - 20,0%). Из жуков наиболее многочисленными являются стафилиниды, щелкуны, долгоносики и жужелицы (рис. 4).
 |
| Рис. 3. Соотношение (%) основных групп почвенных беспозвоночных в сосняках |
Примечание: 1 - дождевые черви; 2 - моллюски; 3 - мокрицы; 4 - паукообразные; 5 - губоногие многоножки; 6 - двупарноногие многоножки; 7 - насекомые;
сокращения типов леса смотри в таблице 1
 |
| Рис. 4. Соотношение (%) семейств жесткокрылых в почвах сосняков |
Примечание: 1 - жужелицы; 2 - стафилиниды; 3 - щелкуны; 4 - долгоносики; 5 - пластинчатоусые; 6 - прочие;
сокращения типов леса смотри в таблице 1
В сосновых лесах Беловежской пущи выявлено 3 типа, 6 классов, 16 отрядов почвенных беспозвоночных. Из них выделены модельные группы - индикаторы почвенных и фитоценотических условий - дождевые черви, жуки-щелкуны, двукрылые, среди которых отмечено 56 видов, относящихся к 40 родам (табл. 4). Основу фаунистических группировок сосняков лишайникового и мшистого составляют ксерофитные (Ampedus balteatus L., Athous haemorrhoidalis F., Prosternon tesselatum L., Sericus brunneus L., Selatosomus aeneus L., Eutolmus rufibarbis Mg., Lasiopogon cinctus Full., Tipula truncorum Meig.) и эврибионтные виды (Dendrobaena octaedra Sav., Athous subfuscus Mull., Dalopius marginatus L., Rhadiurgus variabilis Zett., Thereva sp. и другие). В сосняках кисличных наряду с широко распространенными видами зарегистрированы мезофиты Ectinus aterrimus L., Tipula scripta Meig., T. rubripes Schumm., T. hortulana Meig., Bibio sp., Rhagio sp., Dolichopus sp., Neurogona sp., Phaonia sp. Фауна сосняков черничных представлена мезо-гигрофитами Selatosomus impressus F., Ampedus nigrinus Hbst., Tipula irrorata Macq., а также эврибионтными двукрылыми из семейств Tipulidae, Rhagionidae, Therevidae, Empididae, Asilidae. В сосняке долгомошном зарегистрированы гигрофиты Actenicerus sjaelandicus Mull., Tipula signata Staeg., Smittia sp., Rhamphomyia sp.
Таблица 4
Видовой состав почвенных беспозвоночных в лесах Беловежской пущи
| Виды | Лесная формация | |||||
| сосновая | еловая | дубовая | грабовая | ясеневая | березовая | |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| Lumbricidae | ||||||
| Dendrobaena octaedra Sav. | + | + | + | + | + | + |
| Dendrodrilus rubidus Sav. f. tenuis Eis. | + | + | + | + | + | + |
| Lumbricus rubellus Hoff | + | + | + | + | + | |
| Nicodrilus roseus Sav. | + | + | + | + | + | |
| N. caliginosus Sav. | + | + | + | + | ||
| Octolasium lacteum Oer. | + | + | ||||
| Elateridae | ||||||
| Actenicerus sjaelandicus Mull. | + | |||||
| Ampedus balteatus L. | + | |||||
| A. nigrinus Hbst. | + | + | ||||
| A. sp. | + | |||||
| Athous subfuscus Mull. | + | + | + | + | + | + |
| A. haemorrhoidalis F. | + | + | ||||
| A. vittatus F. | + | + | ||||
| Cardiophorus sp. | + | |||||
| Cydnopus aeruginosus Ol. | + | |||||
| Dalopius marginatus L. | + | + | + | + | + | + |
| Denticollis linearis L. | + | |||||
| Ectinus aterrimus L. | + | + | + | + | + | + |
| Melanotus rufipes Hbst. | + | |||||
| Prosternon tesselatum L. | + | |||||
| Selatosomus aeneus L. | + | + | ||||
| S. impressus F. | + | + | ||||
| S. cruciatus L. | + | + | ||||
| Sericus brunneus L. | + | |||||
| Diptera | ||||||
| Tipulidae | ||||||
| Tipula scripta Meig. | + | + | + | + | + | + |
| T. hortulana Meig. | + | + | + | + | + | |
| T. rubripes Schumm | + | + | + | + | ||
| T. flavolineata Meig. | + | |||||
| T. nubeculosa Meig. | + | + | + | |||
| T. brevispina Pier. | + | + | + | |||
| T. livida V.d.Wulp. | + | + | ||||
| T. irrorata Macq. | + | + | + | |||
| T. lunata L. | + | + | + | |||
| T. variipenis Meig. | + | + | + | |||
| Продолжение таблицы 4 | ||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| T. unca Wied. | + | |||||
| T. signata Staeg. | + | |||||
| T. truncorum Meig. | + | |||||
| T. staegeri Niels. | + | |||||
| T. sp. | + | + | + | |||
| Nephrotoma analis Schumm. | + | + | + | + | ||
| N. flavescens L. | + | |||||
| N. sp. | + | |||||
| Limoniidae | ||||||
| Austrolimnophila sp. | + | |||||
| Hexatominae | + | |||||
| Epiphragma sp. | + | |||||
| Limonia sp. | + | + | ||||
| Pilaria sp. | + | |||||
| Pedicia sp. | + | |||||
| Trimicra sp. (Ormosia sp.) | + | |||||
| Limoniidae gen.sp. | + | + | + | |||
| Ceratopogonidae | + | |||||
| Forcipomyia sp. | + | |||||
| Ceratopogonidae gen.sp. | + | + | + | |||
| Chironomidae | ||||||
| Gymnometriocnemus sp. | + | |||||
| Smittia sp. | + | |||||
| Cecidomyidae | + | |||||
| Monardia sp. | + | |||||
| Bolithophilidae | + | |||||
| Bolithophila sp. | + | |||||
| Mycetophilidae | + | + | + | |||
| Sciaridae | + | + | + | + | + | |
| Bibionidae | ||||||
| Bibio sp. | + | + | + | + | + | + |
| B. marci L. | + | |||||
| Anisopodidae | ||||||
| Silvicola fenestralis Scop. | + | |||||
| Rhagionidae | ||||||
| Chrysopilus sp. | + | + | + | + | + | + |
| Rhagio sp. | + | + | + | + | + | + |
| Symphoromyia sp. | + | + | + | |||
| Xylophagidae | ||||||
| Coenomyia ferruginea Scop. | + | + | + | + | + | |
| Окончание таблицы 4 | ||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 |
| Stratiomyidae | ||||||
| Beris sp. | + | + | ||||
| Microchrysa sp. | + | |||||
| Tabanidae | ||||||
| Haematopoda sp. | + | + | ||||
| Tabanus sp. | + | |||||
| Asilidae | ||||||
| Asilus crabroniformis L. | + | + | + | + | + | |
| Dioctria hyalipennis F. | + | + | + | |||
| D. rufipes De Geer. | + | + | + | |||
| Dismachus sp. | + | |||||
| Eutolmus rufibarbis Mg. | + | |||||
| Lasiopogon cinctus Fall. | + | |||||
| Rhadiurgus variabilis Zett. | + | + | + | + | + | |
| Therevidae | ||||||
| Thereva sp. | + | + | + | + | + | + |
| Empididae | ||||||
| Hilara sp. (Microphorus sp.) | + | + | + | + | + | |
| Ocydromia sp. | + | + | + | + | + | |
| Rhamphomyia sp. (Empis sp.) | + | + | + | |||
| Empididae gen. sp. | + | + | + | + | ||
| Dolichopodidae | ||||||
| Dolichopus sp. | + | + | + | + | + | + |
| Neurogona sp. | + | + | + | + | + | |
| Systenus sp. | + | + | + | + | ||
| Dolichopodidae gen. sp. | + | + | + | |||
| Lauxaniidae (?) | + | + | + | |||
| Phoridae | + | |||||
| Syrphidae | ||||||
| Baccha sp. | + | + | + | + | + | |
| Chrysotoxum sp. | + | + | ||||
| Muscidae | ||||||
| Fannia sp. | + | + | + | + | + | |
| Phaonia sp. | + | + | + | + | + | + |
| Sarcophagidae | + | |||||
Для сосняков характерна низкая плотность дождевых червей (табл. 2, 3), играющих огромное значение в переработке растительного опада и формировании гумусового слоя почвы (Зражевский, 1957). При этом они представлены практически единственным видом D. octaedra (табл. 4), имеющим широкое распространение и населяющим самые разнообразные почвы (Перель, 1979). Даже в сосняке грабово-кисличном, имеющем в составе подстилки большое количество лиственного опада, наблюдается полное доминирование D. octaedra. Лишь единично отмечены подстилочный Dendrodrilus rubidus Sav. f. tenuis Eis. и почвенно-подстилочный вид Lumbricus rubellus Hoff. Для сосняков характерно отсутствие в люмбрицидокомплексах видов дождевых червей, связанных с собственно почвенными слоями, хотя в зоне смешанных лесов в сложных сосняках эти виды Lumbricidae являются доминирующими по плотности и зоомассе (Перель, 1962; Алейникова, 1968).
Своеобразно распределение в различных типах сосняков сапротрофных беспозвоночных, нуждающихся для инкрустации покровов известью в биогенном кальции, - двупарноногих многоножек, моллюсков и мокриц. В сосняках кисличных плотность Diplopoda составляет всего 0,2 - 3,7 экз/м2 (табл. 2, 3). В средневозрастном сосняке черничном их плотность увеличивается до 19,0 экз/м2, а в сосняке долгомошном она достигает величины 92,5 экз/м2. Подобных высоких значений плотности Diplopoda в сосняках с полным преобладанием в составе древостоя сосны в литературе не отмечено. В сосняке долгомошном высока плотность Mollusca (11,2 экз/м2). При плотности 3,5 экз/м2 здесь зарегистрированы мокрицы. Объяснение этого явления, по-видимому, заключается в особенностях формирования почвообразующих пород на территории Беловежской пущи и связанной с этим спецификой почвообразовательных процессов. Неглубокое залегание моренных суглинков обогащает грунтовые воды кальцием и магнием. В результате этого количество СаО (10,5 - 21,0 мг/100 г) в верхних почвенных слоях сосняка долгомошного не уступает таковому сосняков кисличных (7,0 - 33,6 мг/100 г). Наличие в составе древостоя сосняков черничного и долгомошного березы, опад которой содержит большое количество этого элемента и является одним из наиболее предпочитаемых в питании Diplopoda, Mollusca, Oniscoidea (Dunger, 1958), способствует увеличению численности их популяций.
Подавляющее большинство беспозвоночных в сосняках Беловежской пущи сосредоточено в подстилке и самом верхнем минеральном слое (рис. 5), что связано с легким механическим составом дерново-подзолистых почв и маломощностью аккумулятивного горизонта А1. Минеральные горизонты почвы населены лишь личинками жесткокрылых (Curculionidae, Elateridae, Scarabaeidae) и двукрылых (больше Asilidae). Прослеживается тенденция перехода обитания беспозвоночных в подстилку с увеличением ее мощности и влажности почвы. В сосняке долгомошном практически все почвенное население сосредоточено в подстилочном ярусе, что связано с высокой влажностью почвы этого типа леса.
 |
| Рис. 5. Вертикальное распределение (%) почвенных беспозвоночных в сосняках |
Примечание: 1 — подстилка; 2 — 0 - 10 см; 3 — 11 - 20 см; 4 — 21 - 30 см; 5 — 31 - 40 см;
сокращения типов леса смотри в таблице 1
В целом, почвы сосняков Беловежской пущи населены крупными беспозвоночными наиболее бедно в сравнении с другими лесами. Oснову сообществ беспозвоночных сосновых лесов составляют мелкие виды, в результате чего общая зоомасса почвенной мезофауны сосняков довольно низка. Сравнение сообществ почвенных беспозвоночных сосняков Беловежской пущи с сообществами беспозвоночных аналогичных лесов Беларуси и запада России показывает значительное сходство в составе и соотношении массовых групп и доминирующих видов.
В ельниках найдено 3 типа, 6 классов, 15 отрядов почвенных беспозвоночных. Среди модельных индикаторных групп отмечено 37 видов, относящихся к 28 родам (табл. 4). В изученных ельниках встречаются как виды, свойственные таежной зоне (A. subfuscus, S. impressus, T. irrorata и другие), так и виды, обычные в зоне смешанных и широколиственных лесов (D. marginatus, E. aterrimus, A. haemorrhoidalis, T. rubripes, Coenomyia ferruginea Scop., Dioctria hyalipennis F., Chrysopilus sp. и другие). Однако удельный вес последних здесь значительно меньше, чем в еловых лесах зоны смешанных лесов, что отражает специфику зонального и географического положения Беловежской пущи.
Общая плотность почвенных беспозвоночных в различных типах ельников Беловежской пущи колеблется от 376,0 до 443,0 экз/м2 (табл. 5). Общая зоомасса беспозвоночных составляет 3,10 - 6,00 г/м2 (рис. 6), что несколько больше приводимых в литературе значений зоомассы почвенной мезофауны для чистых ельников зоны южной тайги и значительно превышает данные показатели для зон северной и средней тайги. В то же время зоомасса беспозвоночных чистых ельников Беловежской пущи уступает таковой ельников неморальных или ельников со значительной примесью лиственных пород (Яворницкий, 1986). Прослеживается тенденция уменьшения плотности и зоомассы почвенной мезофауны в эдафо-фитоценотическом ряду ельников с увеличением влажности почвы. Максимальные общая плотность почвенных беспозвоночных зарегистрирована в ельнике кисличном (434,0 экз/м2), а общая зоомасса - в ельнике мшистом (6,00 г/м2).
Таблица 5
Средняя плотность (экз/м2) почвенных беспозвоночных ельников
| Группы беспозвоночных |
Ельник лишайниковый |
Ельник мшистый |
Ельник черничный |
| Lumbricidae | 6,0±1,2 | 10,5±1,8 | 4,5±1,4 |
| Mollusca | 2,7±0,9 | 2,2±0,9 | - |
| Oniscoidea | + | - | - |
| Arachnida | 165,0±15,4 | 142,2±12,7 | 136,5±14,5 |
| Chilopoda | 18,8±2,6 | 13,3±2,7 | + |
| Diplopoda | 9,0±1,8 | 1,8±0 8 | + |
| Insecta | 232,0±14,6 | 273,0±14,5 | 234,0±14,7 |
| Hemiptera | 2,2±0,9 | 7,2±1,6 | 1,0±0,5 |
| Dermaptera | 5,5±4,2 | 4,8±2,0 | - |
| Psocoptera | + | + | 1,5±0,7 |
| Neuroptera | + | + | + |
| Homoptera | + | 2,3±0,8 | 1,0±0,6 |
| Blattoptera | 2,0±0,8 | 1,2±0,6 | 2,2±1,0 |
| Coleoptera | 159,0±11,6 | 186,3±11,4 | 183,5±13,1 |
| Carabidae | 10,2±1,8 | 8,0±1,4 | 6,0±1,4 |
| Staphylinidae | 80,5±8,1 | 101,8±8,3 | 93,3±8,8 |
| Elateridae | 38,5±4,4 | 47,5±4,4 | 69,5±7,1 |
| Curculionidae | 13,5±2,3 | 17,8±3,4 | 6,5±2,0 |
| Scarabaeidae | 5,5±2,6 | 1,0±0,5 | 1,2±0,6 |
| Прочие Coleoptera | 10,8±2,1 | 10,2±1,8 | 7,0±1,5 |
| Lepidoptera | 4,5±1,8 | 4,2±1,1 | 4,5±1,2 |
| Hymenoptera | 15,8±2,8 | 7,0±1,5 | 8,0±1,5 |
| Diptera | 47,2±7,0 | 58,2±6,5 | 27,3±4,8 |
| Общая плотность | 434,0±23,0 | 443,0±20,6 | 376,0±24,5 |
Примечание: обозначения смотри в таблице 2
 |
| Рис. 6. Зоомасса (г/м2) основных групп почвенных беспозвоночных в ельниках |
Примечание: обозначения смотри на рисунке 1, сокращения - в таблице 1
В сообществах почвенной мезофауны ельников Беловежской пущи преобладают насекомые (232,0 - 273,0 экз/м2) и паукообразные (136,5 - 165,0 экз/м2) (табл. 5), которые составляют соответственно 53,5 - 62,2% и 32,1 - 38,0% от общей плотности почвенных беспозвоночных (рис. 7). Среди насекомых доминируют жесткокрылые (159,0 - 186,3 экз/м2) и двукрылые (27,3 - 58,2 экз/м2), что составляет для Coleoptera 68,2 - 78,4% и для Diptera 11,7 - 21,3% от общей плотности насекомых. Из жуков наиболее многочисленными являются стафилиниды, щелкуны, долгоносики и жужелицы (рис 8). Вышеназванные группы преобладают и по зоомассе. Такой порядок доминирования характерен для ельников южной тайги.
 |
| Рис. 7. Соотношение (%) основных групп почвенных беспозвоночных в ельниках |
Примечание: обозначения смотри на рисунке 3, сокращения - в таблице 1
 |
| Рис. 8. Соотношение (%) семейств жесткокрылых в почвах ельников |
Примечание: обозначения смотри на рисунке 3, сокращения - в таблице 1
В еловых лесах плотность основных групп сапрофагов также невысока. Максимальная плотность дождевых червей зарегистрирована в ельнике кисличном (10,5 экз/м2) (табл. 5). Среди Lumbricidae полностью господствует D. octaedra, в почве ельника кисличного единично встречаются L. rubellus и Nicodrilus roseus Sav. (табл. 4). Плотность двупарноногих многоножек также невысока, хотя в ельнике мшистом плотность Diplopoda составила 9,0 экз/м2, а зоомасса - 0,52 г/м2 (в ельнике мшистом в составе древостоя несколько больше участие лиственных пород, опад которых охотно используется в пищу двупарноногими многоножками) (табл. 5, рис. 5). В ельнике черничном создаются крайне неблагоприятные условия для жизнедеятельности сапрофагов (высокая кислотность почвы, оторфованность подстилки и другие) и в Беловежской пуще ельники черничные представляют собой типичные подтаежные биогеоценозы, населенные низкопродуктивными бореальными комплексами почвенных беспозвоночных.
В ельниках, как и в сосняках, подавляющее большинство почвенных беспозвоночных сосредоточено в подстилке и верхнем 10 см слое почвы (рис. 9). Однако для ельников характерна большая заселенность почвообитающими животными верхних минеральных горизонтов почвы. В еловых лесах в подстилке сосредоточено менее 50% зоомассы почвенной мезофауны. Характерно сходство в распределении беспозвоночных по почвенному профилю в различных типах ельников.
 |
| Рис. 9. Вертикальное распределение (%) почвенных беспозвоночных в ельниках |
Примечание: обозначения смотри на рисунке 5, сокращения - в таблице 1
В целом, ельники Беловежской пущи заселены беспозвоночными несколько обильнее, чем сосняки. Почвенная фауна ельников Беловежской пущи богаче и разнообразнее, чем в таежной зоне, но структура доминирования сообществ беспозвоночных аналогична таковой для подзоны южной тайги.
В лиственных лесах обнаружено 3 типа, 6 классов, 16 отрядов почвенных беспозвоночных. Среди модельных индикаторных групп выявлен 71 вид, относящийся к 44 родам. В состав фаунистических группировок входят эврибионтные и мезофитные виды почвенных беспозвоночных, обычные для зоны смешанных и широколиственных лесов. В лиственных лесах Беловежской пущи встречаются виды беспозвоночных, наиболее многочисленные в лесостепной зоне - D. marginatus, Athous vittatus F., Coenomyia ferruginea, Tipula nubeculosa Meig., Silvicola fenestralis Scop., Chrysopilus sp. и другие. Одновременно с ними высокой численности достигают A. subfuscus и D. octaedra, доминирующие в таежной зоне. Наибольшее количество видов (54) зарегистрировано в дубравах.
Максимальная плотность беспозвоночных выявлена в почвах дубовых лесов (387,0 - 401,2 экз/м2), минимальная - в почвах березовых лесов (193,0 - 237,8 экз/м2) (табл. 6). Лиственные леса Беловежской пущи в сравнении с южнее и восточнее расположенными районами имеют гораздо более низкую плотность сапрофагов. Плотность дождевых червей в исследованных лесах колеблется от 30,0 до 71,2 экз/м2, а в дубраве с преобладанием дуба скального их плотность составляла всего 12,8 экз/м2. Как и в хвойных лесах, в составе люмбрицидокомплексов доминировал D. octaedra. Плотность двупарноногих многоножек также невысока: от 0,2 - 9,8 экз/м2 в дубовых, грабовых, елово-березовых лесах до 13,0 - 18,8 экз/м2 - в ясеневых и грабово-березовых. Малочисленны в лиственных лесах (за исключением ясенника) моллюски и мокрицы. В почвах лиственных лесов Беловежской пущи большого видового разнообразия и плотности достигают двукрылые, участвующие в разложении листового опада - Tipulidae, Limoniidae, Bibionidae, Muscidae (cм. табл. 4). Особенно многочисленны толстоножки рода Bibio L. Так, в дубраве с преобладанием дуба скального плотность двукрылых Bibio sp. в октябре достигала 862 экз/м2 при зоомассе 16,6 г/м2, что составляло 70% от общей зоомассы почвенных беспозвоночных этого периода. Плотность Bibionidae в дубраве с дубом черешчатым равнялась 1151 особям на 1 м2 (при весе - 7,0 г/м2; 32,7% от общей зоомассы мезофауны). Однако следует отметить, что Bibionidae являются довольно непостоянным компонентом в сообществах почвенных животных. Амплитуда колебания плотности этих двукрылых в разные годы может быть довольно значительной.
Таблица 6
Средняя плотность (экз/м2) почвенных беспозвоночных лиственных лесов
| Группы беспозвоночных |
Дубрава кисличная (дуб скальный) | Дубрава кисличная (дуб черешчатый) | Грабняк кисличный | Ясенник кисличный | Березняк грабово-кисличный | Березняк елово-кисличный |
| Lumbricidae | 12,8±1,8 | 59,9±5,3 | 71,2±6,5 | 33,8±3,9 | 42,2±4,5 | 30,0±3,8 |
| Mollusca | 7,5±1,8 | 2,1±0,6 | 2,8±0,8 | 40,2±5,7 | 8,0±1,5 | 4,5±1,2 |
| Oniscoidea | 1,2±0,8 | – | – | 5,0±1,7 | 3,3±1,0 | – |
| Arachnida | 81,5±9,3 | 65,6±4,8 | 74,0±7,7 | 75,5±6,8 | 23,5±3,7 | 31,7±3,9 |
| Chilopoda | 12,0±2,0 | 3,2±0,8 | 7,0±1,4 | 2,5±1,0 | 6,5±1,4 | 6,2±1,4 |
| Diplopoda | 9,8±3,0 | 1,9±0,6 | + | 13,0±3,0 | 18,8±4,0 | 8,8±2,3 |
| Insecta | 262,2±16,7 | 268,3±12,6 | 193,8±9,1 | 136,5±9,9 | 135,5±16,8 | 111,8±7,2 |
| Hemiptera | 1,0±0,5 | 1,8±0,6 | 8,8±1,8 | 2,5±1,0 | 8,8±1,7 | 8,2±1,8 |
| Dermaptera | 12,0±2,7 | 8,2±2,5 | 22,2±3,5 | + | + | 4,8±1,2 |
| Psocoptera | 14,0±3,9 | 1,6±0,5 | 1,5±0,6 | + | – | – |
| Neuroptera | – | – | + | – | + | – |
| Rhaphidioptera | + | – | – | – | – | – |
| Homoptera | + | 1,6±0,6 | 1,8±0,9 | 2,5±0,8 | + | 1,5±0,6 |
| Blattoptera | – | – | 1,8±1,0 | – | – | – |
| Orthoptera | + | – | – | + | – | – |
| Coleoptera | 161,0±10,5 | 154,5±8,2 | 111,2±6,9 | 90,0±7,8 | 95,2±14,5 | 51,0±3,7 |
| Carabidae | 6,8±1,3 | 10,7±1,6 | 11,2±2,1 | 18,7±2,4 | 7,0±1,3 | 4,5±1,5 |
| Staphylinidae | 100,5±8,1 | 100,6±6,8 | 60,3±5,5 | 58,0±4,1 | 25,0±3,2 | 23,2±2,6 |
| Elateridae | 25,2±3,7 | 11,4±1,7 | 8,2±2,0 | 3,5±0,9 | 10,2±2,0 | 8,8±1,6 |
| Curculionidae | 22,0±2,8 | 24,7±2,8 | 17,5±2,9 | + | 6,5±1,6 | 7,3±1,6 |
| Scarabaeidae | 3,2±2,5 | 3,6±0,9 | 6,0±1,5 | 1,0±0,6 | 43,2±13,3 | 4,2±2,0 |
| Прочие Coleoptera | 3,3±0,8 | 3,5±1,0 | 8,0±1,7 | 8,3±1,6 | 3,3±0,9 | 3,0±1,0 |
| Lepidoptera | 1,0±0,5 | 1,4±0,5 | 1,0±0,5 | 1,2±0,5 | 1,2±0,5 | 1,0±0,5 |
| Hymenoptera | 7,7±3,8 | 3,2±0,8 | 5,0±1,1 | + | + | 3,0±0,8 |
| Diptera | 64,5±8,5 | 96,0±6,7 | 40,0±4,0 | 38,5±4,9 | 28,2±4,4 | 42,3±4,7 |
| Общая плотность | 387,0±23,8 | 401,2±14,6 | 349,0±14,6 | 307,0±21,6 | 237,8±19,8 | 193,0±10,7 |
Почвы, на которых произрастают лиственные леса, характеризуются наиболее высокой общей зоомассой мезофауны (до 16,8 г/м2) (рис. 10). Величина зоомассы почвенной мезофауны в дубравах, ясенниках и березняках с участием ели (не превышающая 7 г/м2) более характерна для дерново-подзолистых почв таежной зоны, чем для зоны смешанных лесов. Вместе с тем грабовые, грабово-березовые и грабово-дубовые леса характеризуются довольно высокой общей зоомассой сообществ почвенных беспозвоночных, достигающей 17 г/м2.
 |
| Рис. 10. Зоомасса (г/м2) основных групп почвенных беспозвоночных в лиственных лесах |
Примечание: обозначения смотри на рисунке 1, сокращения - в таблице 1
В лиственных лесах Беловежской пущи среди беспозвоночных преобладают насекомые (44,5 - 67,8%), паукообразные (9,9 - 24,6%) и дождевые черви (3,3 - 17,8%) (рис. 11). По зоомассе высока доля участия дождевых червей (23,9 - 75,4%). Из насекомых наиболее многочисленными являются жесткокрылые (45,6 - 70,2%) и двукрылые (20,6 - 37,8%). Из жесткокрылых наибольшей плотности и зоомассы достигают стафилиниды, щелкуны, долгоносики, пластинчатоусые и жужелицы (рис. 12). Структура доминирования сообществ почвенных беспозвоночных этих лесов типична для лиственных фитоценозов зоны смешанных и широколиственных лесов.
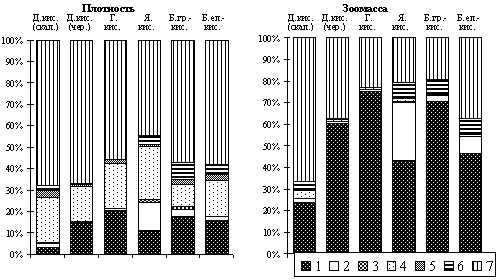 |
| Рис. 11. Соотношение (%) основных групп почвенных беспозвоночных в лиственных лесах |
Примечание: обозначения смотри на рисунке 3, сокращения - в таблице 1
 |
| Рис. 12. Соотношение (%) семейств жесткокрылых в почвах лиственных лесов |
Примечание: обозначения смотри на рисунке 4, сокращения - в таблице 1
Одной из особенностей почвенной фауны лесов Беловежской пущи является то, что даже в лиственных типах леса почвенное население сосредоточено преимущественно в подстилке и самом верхнем минеральном слое почвы (рис. 13). Это обусловлено в первую очередь малочисленностью или отсутствием почвенных видов дождевых червей. В распределении зоомассы почвенный мезофауны лиственных лесов отмечается несколько большая ее "погруженность" в сравнении с хвойными типами леса, связанная с более высокой численностью крупных видов беспозвоночных, обитающих в толще почвы.
 |
| Рис. 13. Вертикальное распределение (%) почвенных беспозвоночных в лиственных лесах |
Примечание: обозначения смотри на рисунке 5, сокращения - в таблице 1
Таким образом, в составе, структурной организации и распределении сообществ почвенных беспозвоночных лиственных лесов Беловежской пущи наблюдается большое сходство с почвенной мезофауной умеренно увлажненных лесов, произрастающих на дерново-подзолистых почвах. Почвенно-зоологические исследования показали, что бурые лесные почвы под дубовыми лесами в Беловежской пуще слабо населены крупными беспозвоночными и, особенно, сапрофагами. Группировки почвенных беспозвоночных, населяющих буроземы Беловежской пущи, представляют собой обедненные аналоги почвенных сообществ широколиственных лесов Средней Европы.
В лесах Беловежской пущи выявлено 3 типа, 6 классов, 17 отрядов почвенных беспозвоночных. Из них выделены модельные группы - индикаторы почвенных и фитоценотических условий - дождевые черви, жуки-щелкуны, двукрылые, среди которых отмечено 94 вида, относящихся к 58 родам, 23 семействам. Основу фаунистических группировок в лесах Беловежской пущи составляют виды, обычные для зоны смешанных лесов. Однако географическое положение пущи на стыке двух лесорастительных областей, а также особенности климата, обеспечивают проникновение на ее территорию как бореальных, так и неморальных элементов фауны. Здесь найдены распространенные в южных широколиственных лесах щелкуны E. aterrimus, A. haemorrhoidalis, A. vittatus, S. cruciatus, двукрылые C. ferruginea, T. nubeculosa и другие. Высокой плотности достигает щелкун D. marginatus. Неморальные элементы в Беловежской пуще приурочены в основном к дубово-грабовым лесам, однако они могут заселять и сосново-еловые фитоценозы. Полесье в периоды последнего материкового оледенения представляло собой перигляциальную зону, что способствовало широкому распространению в Беловежской пуще таких бореальных видов, как A. subfuscus, A. balteatus, S. impressus, T. irrorata и других.
Почвенное население лесов Беловежской пущи по характеру распространения существенно отличается от такового в типичных зональных лесных районах. Холмистый рельеф, пестрота почвенного и растительного покрова, сочетание бореальных и неморальных растительных элементов в фитоценозах пущи определяют многообразие различных по составу и структуре группировок беспозвоночных. В пуще одновременно с обычными для зоны смешанных лесов Восточной и Средней Европы комплексами беспозвоночных животных, населяющими дерново-подзолистые почвы хвойных и смешанных лесов, широко распространены типично бореальные сообщества, характерные для ельников южной тайги. Вместе с ними в пуще встречаются неморальные сообщества почвенных беспозвоночных. Они приурочены к грабовым, дубовым и березовым лесам и по составу населения во многом схожи с аналогичными группировками подзоны широколиственных лесов.
Основная масса почвенного населения в лесах пущи представлена поверхностно живущими видами или же видами, обитающими в самых верхних почвенных горизонтах (в среднем до 95%). Распространение собственно почвенных видов даже в более плодородных почвах лиственных лесов ограничено.
Анализ материала по методу Маунтфорда на основе коэффициента Жаккара четко показывает, что распределение комплексов беспозвоночных тесно связано с гидро-термическими условиями почвы, по характеру которых обследованные типы леса можно разделить на 4 группы: 1) сухие леса (сосняки лишайниковый и мшистые); 2) свежие (ельник мшистый и кисличные типы леса); 3) сырые (сосняки и ельник черничные); 4) заболоченные (сосняк долгомошный) (рис. 14). При этом иллюстративное построение диаграммы наглядно показывает, насколько точно совпадает расположение фаунистических группировок беспозвоночных с расположением типов леса в лесоводческой классификации (при построении ее по градиенту увеличения влажности почвы) (Юркевич, 1980). Степень сходства комплексов беспозвоночных, рассчитанная по методу Маунтфорда на основе коэффициента Наумова, характеризует связь беспозвоночных с типом гумусообразования. По характеру образующегося гумуса все обследованные почвы делятся на: 1) муллевые (ясенник кисличный); 2) модеровые (лиственные типы леса); 3) моровые (хвойные типы леса); 4) моровые с элементами торфонакопления (сосняк долгомошный). Особняком выделяется почва сосняка лишайникового, имеющего второй погруженный гумусовый горизонт. В данном распределении также обнаруживается высокая точность совпадения расположения отдельных комплексов беспозвоночных с расположением типов почв в почвоведческой классификации (при построении ее по градиенту увеличения доли мягкого гумуса) (Почвоведение, 1975).
 |
| Рис. 14. Степень сходства (%) между комплексами почвенных беспозвоночных лесов Беловежской пущи (I - расчет по коэффициенту Жаккара; II - расчет по коэффициенту Наумова) |
Примечание: 1 - Сосняк лишайниковый; 2 - С. мшистый (60 лет); 3 - С. черничный (60 лет); 4 - С. долгомошный; 5 - С .мшистый (150 лет); 6 - С. елово-кисличный; 7 - С. грабово-кисличный; 8 - С. черничный (150 лет); 9 - Ельник мшистый; 10 - Е. кисличный; 11 - Е. черничный; 12 - Дубрава кисличная (дуб скальный); 13 - Д .кисличная (дуб черешчатый); 14 - Д. кисличная (контрольная площадь); 15 -Грабняк кисличный; 16 - Ясенник кисличный; 17 - Березняк грабово-кисличный; 18 - Б. елово-кисличный; 19 - Д. кис. (зубропитомник)
Таким образом, распределение почвенных беспозвоночных в лесах Беловежской пущи тесно связано со всем комплексом природных условий (с рельефом, характером и строением почвообразующих пород, гидрологией, составом растительности и другими), ведущими среди которых являются влажность и тип почвы. В схожих почвенных условиях структурная организация сообществ почвенных животных в значительной степени определяется составом древостоя.
1. Алейникова М. М. Почвообитающие беспозвоночные различных типов леса в Среднем Поволжье //Матер. по фауне и экологии почвообитающ. беспозвоночных. - Казань, 1968. - С. 3-97.
2. Гиляров М. С. Методы количественного учета почвенной мезофауны //Почвоведение. - 1941. - №4. - С. 48-77.
3. Зайцев Г. Н. Математическая статистика в экспериментальной ботанике. - М., 1984. - 424 с.
4. Зражевский А. И. Дождевые черви как фактор плодородия лесных почв. - Киев, 1957. - 271 с.
5. Ковальков М. П., Балюк С. С., Будниченко Н. И. Беловежская пуща. Аннотированный указатель отечественной литературы (1935 - 1983 гг.). - Мн., 1985. - 336 с.
6. Количественные методы в почвенной зоологии. - М., 1987. - 288 с.
7. Перель Т. С. Комплексы почвенных беспозвоночных в сложных сосняках различных типов //Pedobiologia. - 1962. - Vol. 1, №3. - Р. 174-188.
8. Перель Т. С. Распространение и закономерности распределения дождевых червей фауны СССР (с определительными таблицами Lumbricidae и других Megadrili). - М., 1979. - 272 с.
9. Почвоведение. - М., 1975. - 496 с.
10. Чернов Ю. И. Основные синэкологические характеристики почвенных беспозвоночных и методы их анализа //Методы почв.-зоол. исслед. - М., 1975. - C. 160-216.
11. Юркевич И.Д. Выделение типов леса при лесоустроительных работах. - Мн., 1980. - 120 с.
12. Яворницкий В. И. Сообщества почвенной мезофауны некоторых экосистем Карпатского государственного национального природного парка //Пробл. охраны генофонда и упр. экосистемами в заповед. лес. зоны. Тез. докл. Всес. совещ., Березинский заповедник, 23-25 сент., 1986. Ч. 2. - М., 1986. - C. 235-237.
13. Dunger W. Uber die Zersetzung der Laubstreu durch die Boden-Makrofauna im Auenwald //Zool. Jahrb. Abt. 3. - 1958. - B. 86, H 1\2.
14. Karpinski J., Okolow Cz. Bibliografia Bialowieska. - Warszawa, 1969. - 208 p.
15. Okolow Cz. Bibliografia Puszczy Bialowieskey 1967 - 1972. - Bialowieza, 1976. - 164 p.
16. Okolow Cz. Bibliografia Puszczy Bialowieskey 1973 - 1980. - Bialowieza, 1983. - 190 p.
Опубликовано на стр. 161 - 182.